|
|
 |
|
Судьба макроконхов. Почему их так мало?
Автор: Alex
Рубрика: Палеонтологические публикации
Все головоногие моллюски раздельнополы – среди них есть как самцы, так и самки и менять пол на протяжении жизни они не могут (хотя среди более примитивных моллюсков такое встречается). Несомненно, раздельнополыми были и аммониты – далеко не примитивные представители головоногих моллюсков. Согласно современной теории полового диморфизма аммонитов, самки были крупнее самцов, их раковины мы называем макроконхами (или мегаконхами – это частный случай макроконхов). Раковины самцов соответственно называются микроконхами (есть еще миниконхи – мелкий частный случай микроконхов). Для многих коллекционеров крупная раковина макроконха – находка куда более интересная и желанная, чем микроконховая мелочь. Но вот проблема – далеко не во всех местонахождениях и не во всех слоях макроконхи встречаются также часто как микроконхи. Зачастую их раковины попадаются существенно реже, а кое-где их и вообще нет, хотя микроконхов в этих же местах может быть очень и очень много. Почему? Неужели иногда самцы жили вообще без самок? Или палеонтологи не правы в своих интерпретациях микро- и макроконхов?
Этот вопрос неоднократно поднимался на "Аммоните.ру", поэтому я решил сделать специальную публикацию по проблеме "отсутствующих макроконхов". Чтобы разобраться в этом вопросе нам, как это часто бывает в палеонтологии, стоит обратить внимание на современных головоногих моллюсков.
Автор теории полового диморфизма аммонитов, Г.Маковский, в своей публикации, посвященной этому вопросу (Makowski 1962) отметил, что численное соотношение диморфов (самцов и самок) аммонитов должно быть соизмеримо с соотношением полов в современных популяциях. Однако современные цефалоподы очень сильно различаются между собой – структура популяции у медленно размножающихся и долго живущих наутилусов кардинально отличается от структуры популяции, например, у кальмаров, успевающих вырасти, оставить потомство и умереть естественной смертью за 1-2 года.
У наутилусов соотношение числа самцов и самок составляет примерно 1:1. Но аммониты по происхождению, многим деталям анатомии и вероятно по продолжительности жизни были ближе к колеоидеям (осьминогам, кальмарам, каракатицам) чем к наутилусам и вряд ли могли иметь наутилусовую структуру популяции. Хотя соотношение самцов и самок 1:1 наверняка встречалось в популяциях аммонитов, оно не было общим правилом.
Ближайшим современным экологическим аналогом аммонитов, по-видимому, являются эпипелагические осьминоги надсемейства Argonautoidea. Их диморфизм очень похож на диморфизм аммонитов – например у видов Argonauta argo и Ocythoe tuberculata средняя длина туловища (точнее, длина мантии) самок составляет 20-30 см, а самцов лишь 2-3 см. Впрочем, у самок Argonauta hians длина туловища составляет лишь 5-7 см (а самцов 1-2 см), так что тут разница в размерах не в 10 раз, как у A.argo, а лишь в два-три раза. Тем не менее, у аргонаутоидей самцы всегда заметно меньше самок. Все самки не откладывают, а вынашивают яйца (как, по всей видимости, это делали и аммониты): самки Argonauta хранят яйца в специальной раковине, Tremoctopus носят в руках, а Ocythoe вообще перешли к живорождению. Все они, так или иначе, пытаются восполнить отсутствие фрагмокона и разрабатывают разные варианты поддержания плавучести: самки Argonauta загоняют пузырек воздуха в раковину, Tremoctopus парят на широких лопастях-перепонках на краях щупалец, Ocythoe имеют плавательный пузырь. Самцы всех этих осьминогов не имеют специальных приспособлений для поддержания плавучести, но они залезают внутрь парящих в воде оболочников – ищут себе замену отсутствующей раковины (Несис 2005).
Данных по соотношению численности самцов и самок у Argonautoidea мне найти не удалось. Но К.Несис (2005) говорит о том, что внутри мантийной полости самок Argonauta и Tremoctopus обычно хранится 2-3 гектокотиля. Гектокотиль – это специальная модифицированная рука самца, содержащая внутри себя сперматофор (пакет со спермой) и передающаяся самке при спаривании. Раньше в книжках писали, что гектокотиль якобы самостоятельно гоняется по морю за удирающей от него самкой как торпеда за подводной лодкой )) Сейчас уже понятно, что это ерунда, но гектокотиль действительно отрывается от тела самца и заползает в мантийное отверстие самки (правда для этого осьминоги должны соприкасаться). Важно то, что самец аргонаутоидей несет лишь один гектокотиль и за всю свою жизнь может спариться лишь один единственный раз (у других колеоидей все устроено немного иначе), после этого он либо умирает самостоятельно, либо самка просто его съедает (вот так не повезло этим животным). Так как неоплодотворенных взрослых самок аргонаутоидей практически не находят, значит, наличие 2-3 гектокотилей практически у каждой самки свидетельствует о сильном перекосе в численности в пользу самцов – на каждую самку должно приходиться по 2-3 самца. О причинах этого можно только догадываться, но возможно, что крупные и фактически беззащитные самки просто чаще становятся добычей хищников, чем мелкие самцы, которых сложнее заметить.
Так, по аналогии с аргонаутоидеями мы можем выдвинуть первую версию, первое объяснение того, почему у аммонитов микроконхов часто оказывается больше, чем макроконхов.
1. Возможно, что в части популяций аммонитов действительно существовал дисбаланс в соотношении числа самцов и самок (самцов могло быть в полтора-три раза больше, например, из-за высокой смертности более крупных и заметных самок).
Возможно, что подобную ситуацию мы наблюдаем в кимериджских отложениях в Липицах у аммонитов семейства Aspidoceratidae. Там микроконхи (Sutneria) встречаются раза в 3 чаще, чем целые макроконхи того же вида (выделенные правда в отдельный род Aspidoceras). При этом довольно часто попадаются некрупные Aspidoceras, которые явно погибли, не достигнув даже половины взрослого размера раковины.

Фото Липицкая мелочь, автор VitalyNcr
Судя по мощным шипам макроконхов аспидоцератид, они находились под серьезным прессом хищников. У микроконхов-сутнерий шипов не было, видимо, их защищал малый размер (эксперименты с современными хищниками показывают, что у них, как правило, есть «нижняя граница размера добычи» - они просто не обращают внимания на мелочь, которую им невыгодно ловить – усилий на поимку надо много, а есть там почти нечего). А вот самок-аспидоцерасов, несмотря на их шипы периодически все-таки ели, и ели довольно активно, поэтому во взрослой популяции мелких самцов оказывалось больше.
Но это лишь один из вариантов, одно из возможных объяснений преобладания микроконхов над макроконхами. У других аммонитов могли быть и другие причины такого феномена. Современные головоногие подсказывают нам и еще одну версию. К.Несис (2005) пишет, что среди современных кальмаров многие виды совершают длительные миграции – сначала молодь плывет к местам, где можно хорошо поесть и быстро вырасти, затем кальмары возвращаются к местам нереста. У тихоокеанского кальмара (Todarodes pacificus) по словам Несиса максимальная протяженность миграции кальмаров зимней группировки от острова Кюсю до восточной части Камчатки - составляет 3500-4000 км. И вот что еще пишет Несис: «Кальмариха может спариваться перед самым нерестом, будучи вполне половозрелой (как у командорского кальмара), а может и задолго до нереста, месяца за два, за три, будучи еще совсем незрелой (как у тихоокеанского). В этом случае самцов на нерестилище вовсе не бывает, их к тому времени, может, уже и на свете-то нет.» После спаривания самцы кальмаров погибают почти сразу, самки погибают после откладки яиц. Выходит, что между точками, где на дно опускаются умершие самцы кальмара, и где покоятся самки, может быть расстояние в несколько тысяч километров. Вряд ли аммониты могли мигрировать на такие расстояния, но десятки или даже пару сотен километров они наверняка могли проплыть. Недавно как раз вышла статья (Yahada, Wani 2013) подтверждающая существование миграций у некоторых аммонитов. У аммонитов, судя по всему, как и у кальмаров с осьминогами, самцы после спаривания погибали. А вот самки оставались – им нужно было выносить яйца или даже мигрировать с ними в район, более подходящий для жизни детенышей (а уже после этого умереть с чувством выполненного долга). Таким образом, второй причиной несоответствия числа микро-и макроконхов может быть следующая:
2. Миграции самок аммонитов к местам нереста (или рождения молоди) и как следствие раздельное захоронение самок и самцов (самцы и захораниваются в районе спаривания, самки – в районе откладки яиц или рождения молоди).
В таком случае могут образовываться раздельные захоронения самцов и самок (при этом на захоронении самцов самки тоже могут изредка встречаться – погибшие от каких-то естественных причин или от хищников). Но теоретически отдельного захоронения самок может вообще и не быть – если они откладывали яйца или рожали живых детенышей в прибрежной зоне, то их раковины могли просто истираться в песок у берегов. Возможно, что с подобной ситуацией мы сталкиваемся в отложениях верхнеоксфордской зоны Amoeboceras alternoides в Марково.

Фото Пришельцы из прошлого., автор Denozavr

Фото Гладкий макроконх Amoeboceras alternoides, автор Alex
Эти самые Amoeboceras alternoides там представлены почти исключительно самцами-микроконхами разного размера, раковины макроконхов крайне редки. Похожая ситуация наблюдается и в Карачаево-Черкесии в долине реки Б. Зеленчук, где микроконхи Parkinsonia встречаются во много раз чаще, чем макроконхи. Соотношение микро-и макроконхов в этих слоях позволяет исключить первый вариант, который мы обсуждали выше (на одну самку никак не могли приходиться сотни самцов – это не рационально), зато вариант с миграцией самок здесь вполне возможен.
Впрочем, здесь может быть и еще одно объяснение. До сих пор мы исходили из того, что находки раковин аммонитов на дне примерно отражают их реальную численность в толще воды. Но ведь это совсем не обязательно! Ведь мы, раскапывая местонахождение, имеем дело не с биоценозом (совокупностью живущих на определенной территории организмов), а с ориктоценозом (совокупностью того, что тут захоронилось и сумело окаменеть). И сохраняется в ископаемом состоянии далеко не все. И опыт показывает (об этом говорил еще Маковский), что в определенных условиях крупные раковины аммонитов сохраняются гораздо хуже. То есть третья причина отсутствия макроконхов это:
3. Плохой «фоссилизационный потенциал» крупных раковин.
Хотя есть любимые всеми палеонтологами местонахождения, в которых крупные аммониты сохранились замечательно (например, в Никитино или Глебово), но есть места, где мы видим отличные примеры исчезновения макроконхов в процессе фоссилизации. Например, это отложения зоны Kachpurites fulgens в Москве и Московской области (Еганово, Мневники, Фили). Средний размер аммонитов из этой зоны – 5-7 см, раковины в 10-12 см в диаметре уже смотрятся гигантами. Но при внимательных раскопках становится понятно, что в слоях наряду с этой мелочью захоронено и множество гораздо более крупных раковин диаметром около 30 см. Только они представляют собой лишь прослои перламутра в глине. Если маленького аммонита, даже размокшего или растрескавшегося, можно как-то вытащить вместе с куском глины и сохранить, то «гнилую» 30-тисантиметровую раковину не то что извлечь невозможно, ее часто не удается даже расчистить для фотографирования и определить хотя бы до рода.
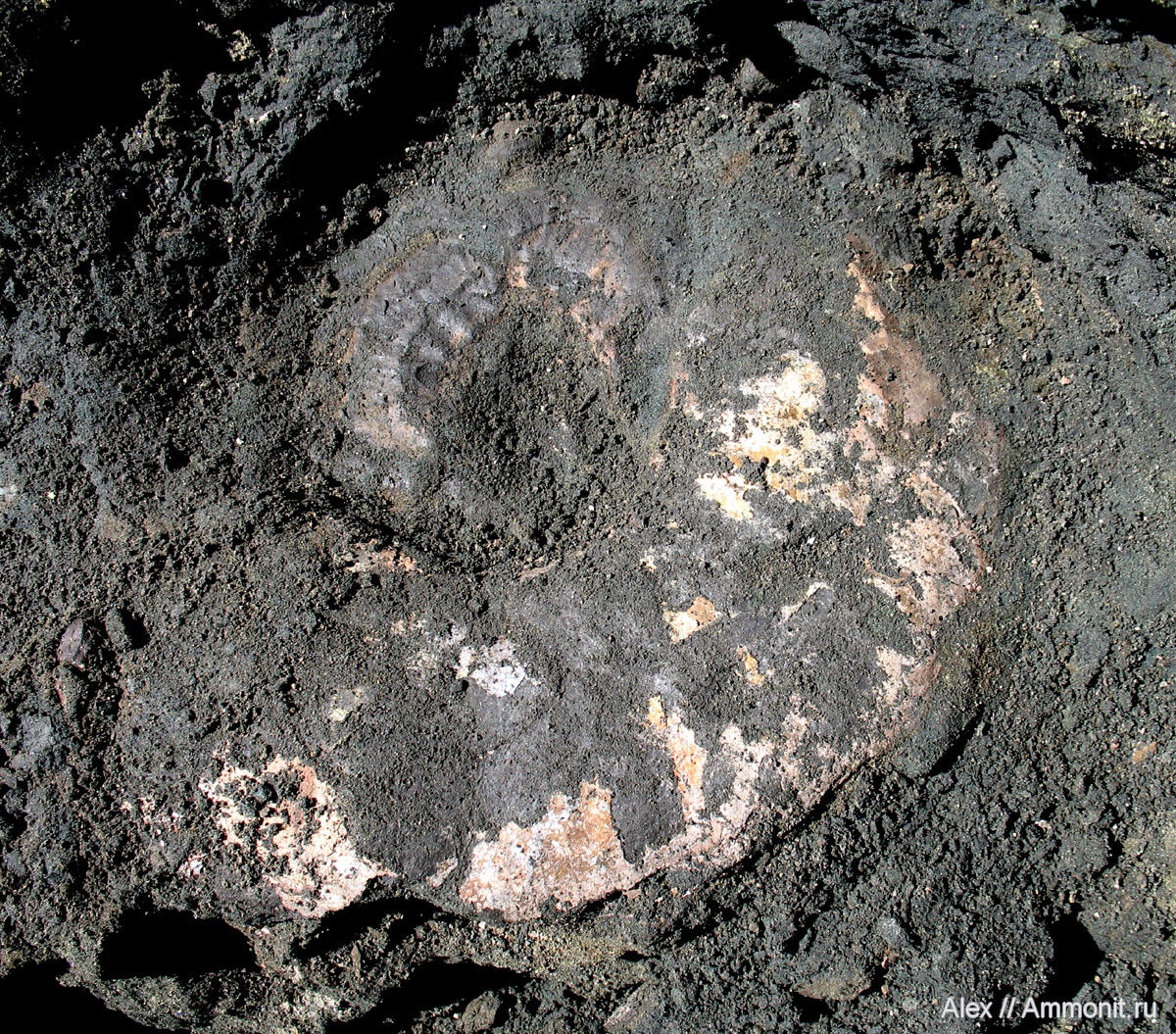
Фото Гигант из зоны fulgens, автор Alex
Причина такой отвратительно сохранности крупных раковин, по-видимому, кроется в сочетании физических и химических факторов. Конкреции, обеспечивающие хорошую сохранность раковин, чаще формируются вокруг небольших объектов или пупков крупных раковин – остальное разрушается. Крупную раковину вероятнее разломают падальщики на дне или волны (если они достают до дна в этом месте), ей сложнее погрузиться в осадок и тем самым защититься от различных сверлильщиков и обрастателей, также способных разрушить раковину. Крупную раковину скорее раздавит вес вышележащих слоев, чем мелкую, размоет просачивающаяся в слой вода. То есть, макроконхи могут исчезать (или почти исчезать) из местонахождения уже после захоронения, а могут разрушаться на дне моря.
В уже упоминавшихся слоях зоны A. alternoides в Марково сохранность макроконхов гораздо хуже, чем микроконхов, значит, там этот фактор действовал. Тем не менее, макроконхи там хотя и раздавленные и разломанные, но не настолько чтобы они могли полностью исчезнуть. Похожая картина и на реке Б. Зеленчук – макроконхи там фрагментарны, но вряд ли они могли исчезать полностью. Даже если мы пропускаем два из трех макроконхов (что вряд ли), то все равно преобладание микроконхов слишком уж огромно. В чем же дело?
На самом деле есть еще одна, четвертая причина исчезновения макроконхов. О ней нам «рассказали» современные наутилусы. Японские исследователи (Wani et al. 2005), изучавшие плавучесть раковин умерших наутилусов обнаружили, что посмертная судьба раковины зависит от ее размера. Маленькая раковина гарантировано быстро утонет, а вот большая, скорее всего, будет держаться в воде куда дольше и даже может всплыть к поверхности. Данный вывод может на первый взгляд противоречить теоретическим предположениям о том, что в большую раковину через крупное отверстие сифона вода зальется легче и быстрее, однако это так, выводы подтверждены экспериментами. Раньше, кстати, писали, что скорость затопления раковин зависит от глубины – чем на большей глубине умер наутилус, тем быстрее он утонет (большее давление воды быстрее выдавит газ из фрагмокона). Но данные японских специалистов не подтвердили это предположение – все оказалось даже наоборот. Эксперимент показал, что затопление раковины начинается только после частичного разложения тела, особенно, участка около сифона. А у поверхности разложение идет быстрее, чем на глубине (так как у поверхности значительно теплее). Скорее всего, точно также – только после частичного разложения мягких тканей, тонули и аммониты. Это, кстати, косвенным образом объясняет отсутствие в палеонтологической летописи отпечатков тел наутилусов и аммонитов – если кальмар или осьминог могут утонуть весьма свежими и сохраниться в донном осадке еще до начала сильного разложения, то наутилусы и аммониты попадают на дно уже весьма подпорченными.
Эксперименты показали, что некоторые крупные раковины наутилусов, которые не утонули быстро, после разложения мягкого тела могут всплыть. Со временем они, может быть, и утонут, но тут многое зависит от гидрологического режима данного места: сильные течения в приповерхностном слое воды могут унести их за сотни километров, а то и прибить к берегу. Для маленьких раковин такой сценарий исключен, а вот для больших – вполне возможен. То есть еще одна причина малочисленности макроконхов это:
4. Разная скорость затопления раковин и как следствие этого избирательный перенос крупных раковин макроконхов в другое место, а то и в прибрежную зону.
Получается та же избирательная миграция раковин что и во втором варианте, только посмертная. Границу между затоплением и всплытием у наутилусов авторы упомянутого исследования проводят по диаметру 20 см. Они утверждают, что эти расчеты полностью применимы и для аммонитов – то есть аммониты диаметром меньше 20 см после смерти тонули, а больше 20 см могли долго оставаться в воде и совершать длительные посмертные путешествия. Но мне кажется, что это слишком уж широкое обобщение. Аммониты очень сильно различались по форме и весу раковин, по длине жилой камеры и объему фрагмокона. Наверняка у разных аммонитов эта граница несколько смещалась в ту или иную сторону. Судя по всему, мощные шипастые аспидоцерасы с тяжелыми раковинами и аптихами и крупные Perisphinctidae обладали неплохой способностью к затоплению почти независимо от размера, чего нельзя сказать о многих других аммонитах.
Но вполне вероятно, что именно эти посмертные миграции были причиной исчезновения макроконхов Amoeboceras из слоев зоны alternoides. Раковины макроконхов могли всплывать и уноситься течениями в неизведанную даль, а микроконхи тонули там, где и жили.
Но почему в следующей зоне – Amoeboceras serratum – вдруг появляется множество амоебоцерасовых макроконхов? Тут может быть сразу четыре варианта объяснения. Если в эпоху alternoides живые макроконхи все-таки куда-то мигрировали, то в эпоху serratum они могли от этого отказаться (например, если пищи для аммонителл стало больше и миграции стали излишними – зачем куда-то плыть, если нас и здесь неплохо кормят? :). Если мигрировали лишь пустые раковины мертвых макроконхов, то могла либо измениться температура или глубина обитания аммонитов (тела стали разлагаться очень быстро и даже крупные раковины быстро тонули), либо уменьшился средний размер самих макроконхов (и они перешагнули эту границу, отделяющую утопление от всплытия), либо изменились гидрологические условия (течения стали слабыми и даже долго болтающиеся в воде раковины не уносило далеко). Как это было на самом деле пока сказать невозможно.
Так что в отсутствии макроконхов (или их малом количестве) во многих местонахождениях нет ничего удивительного. Правда, чтобы понять, какая из четырех причин (а может быть есть и другие) привела к дисбалансу численности половых диморфов в каждом конкретном случае, нужно очень внимательно изучать каждое местонахождение. К тому же надо помнить, что у некоторых видов и микроконхи от макроконхов почти не отличаются, кажущийся дисбаланс может быть связан с неправильной оценкой принадлежности раковин (хотя это довольно экзотическая ситуация). Но иногда все-таки предположить, какой фактор сыграл главную роль. Встретив совместное захоронение микро- и макроконхов с адекватным соотношением численности (от 1:1 до приблизительно 1:4 в пользу микроконхов) можно предположить, что мы имеем дело со «слепком» настоящей популяции. Наблюдая захоронение с преобладанием макроконхов (бывает и такое) можно обратить внимание на их сохранность (мертвые и долго плававшие раковины обычно несут на себе много обрастателей), если раковины чистые и неповрежденные, вряд ли их принесло издалека уже пустыми. Однако, достаточно сложно различить ситуации с миграцией живых макроконхов или посмертным выносом их раковин, если у нас есть лишь «микроконховое захоронение». Но может быть в некоторых случаях и такие загадки удастся со временем разгадать.
Литература:
Makowski H. 1962. Problem of sexual dimorphism in ammonites // Palaeontologica Polonica 1962, № 12. P. 1–92.
Wani R., Kase T., Shigeta Y., De Ocampo R. 2005 New look at ammonoid taphonomy, based on field experiments with modern chambered nautilus
Yahada H., Wani R. 2013. Limited migration of scaphitid ammonoids; evidence from the analyses of shell whorls. Journal of Paleontology. 87(3)
Несис, К. Н. 2005. Головоногие: умные и стремительные: истории из частной и семейнной жизни кальмаров, каракатиц, осьминогов, а также наутилуса помпилиуса. Москва: Октопус, 2005.
Ключевые слова (тэги): аммониты, макроконхи, микроконхи, диморфизм, Ammonites, Microconchs, Macroconchs
 Добавить в избранное
Добавить в избранное
|
Постоянный адрес публикации: |
HTML-ссылка на публикацию: |
Публикация создана 5 августа 2014 года
Если вы хотите оставить комментарий,
вам нужно зарегистрироваться (или авторизоваться, если вы уже зарегистрированы)
Комментарии:
|
<a href="http://www.ammonit.ru/text/1327.htm#196487">http://www.ammonit.ru/text/1327.htm#196487</a> |
|
|||||
А по-моему нельзя говорить об одной конкретной причине - их вполне могло быть несколько. | ||||||
|
комментарий 1 уровня |
Комментарий создан 5 августа 2014 года в 14:14:24 |
|||||
 Добавить в избранное
Добавить в избранное 


|
<a href="http://www.ammonit.ru/text/1327.htm#196496">http://www.ammonit.ru/text/1327.htm#196496</a> |
|
|||||
Так в общем статья о том же самом. В каждом конкретном случае, все-таки, скорее всего была одна причина (хотя могло быть и несколько). | ||||||
|
комментарий 2 уровня |
Комментарий создан 5 августа 2014 года в 17:22:41 |
|||||

|
<a href="http://www.ammonit.ru/text/1327.htm#196492">http://www.ammonit.ru/text/1327.htm#196492</a> |
|
|||||
Проглотил вдумчиво неотрываясь...Есть над чем поразмыслить. Доходчиво и понятно.Спасибо Александр! | ||||||
|
комментарий 1 уровня |
Комментарий создан 5 августа 2014 года в 15:45:50 |
|||||

|
<a href="http://www.ammonit.ru/text/1327.htm#196497">http://www.ammonit.ru/text/1327.htm#196497</a> |
|
|||||
Спасибо! | ||||||
|
комментарий 2 уровня |
Комментарий создан 5 августа 2014 года в 17:24:05 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#196513">http://www.ammonit.ru/text/1327.htm#196513</a> |
|
|||||
Интересная публикация.
| ||||||
|
комментарий 1 уровня |
Комментарий создан 5 августа 2014 года в 19:11:12 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#196520">http://www.ammonit.ru/text/1327.htm#196520</a> |
|
|||||
Вот это интересное наблюдение. В принципе можно предположить, что раз макроконхи одного вида есть, а другого нет, то дело не в пассивном посмертном выносе и не только в плохой сохранности, а в прижизненной миграции макроконхов Amoeboceras. Хотя конечно могут быть и какие-то другие варианты. | ||||||
|
комментарий 2 уровня |
Комментарий создан 5 августа 2014 года в 19:34:07 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#196516">http://www.ammonit.ru/text/1327.htm#196516</a> |
|
|||||
Замечательная публикация!
| ||||||
|
комментарий 1 уровня |
Комментарий создан 5 августа 2014 года в 19:21:33 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#196522">http://www.ammonit.ru/text/1327.htm#196522</a> |
|
|||||
Спасибо! Да, судя по этим фрагментам макроконхи паркинсоний действительно просто плохо сохранились.
| ||||||
|
комментарий 2 уровня |
Комментарий создан 5 августа 2014 года в 19:43:42 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#196523">http://www.ammonit.ru/text/1327.htm#196523</a> |
|
|||||
Можете обратить внимания и на края фрагментов - они окатаны и заглажены. Т.Е. это не свежие и острые изломы от целых раковин, а именно отдельные остатки. которые изначально сохранялись уже обломками.
| ||||||
|
комментарий 3 уровня |
Комментарий создан 5 августа 2014 года в 19:53:16 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#196531">http://www.ammonit.ru/text/1327.htm#196531</a> |
|
|||||
Если они изначально были обломками, то вероятнее вариант что это фрагменты случайно погибших макроконхов, а основное захоронение макроконхов было в другом месте, и они туда мигрировали. | ||||||
|
комментарий 4 уровня |
Комментарий создан 5 августа 2014 года в 21:25:48 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#196536">http://www.ammonit.ru/text/1327.htm#196536</a> |
|
|||||
Может и так. Конечно, если бы найти захоронение макроконхов именно этого рода (Parkinsonia) недалеко от разреза, многие бы вопросы отпали.
| ||||||
|
комментарий 5 уровня |
Комментарий создан 5 августа 2014 года в 22:04:20 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#196568">http://www.ammonit.ru/text/1327.htm#196568</a> |
|
|||||
Интересно-то как! Спасибо за публикацию, Александр! :) | ||||||
|
комментарий 1 уровня |
Комментарий создан 5 августа 2014 года в 23:00:58 |
|||||

|
<a href="http://www.ammonit.ru/text/1327.htm#196856">http://www.ammonit.ru/text/1327.htm#196856</a> |
|
|||||
Очень интересная публикация!
| ||||||
|
комментарий 1 уровня |
Комментарий создан 8 августа 2014 года в 16:03:39 |
|||||

|
<a href="http://www.ammonit.ru/text/1327.htm#197152">http://www.ammonit.ru/text/1327.htm#197152</a> |
|
|||||
Интересно. Спасибо. | ||||||
|
комментарий 1 уровня |
Комментарий создан 11 августа 2014 года в 07:38:52 |
|||||

|
<a href="http://www.ammonit.ru/text/1327.htm#197279">http://www.ammonit.ru/text/1327.htm#197279</a> |
|
|||||
То, о чем хотела узнать, спросить!Спасибо! | ||||||
|
комментарий 1 уровня |
Комментарий создан 11 августа 2014 года в 21:27:49 |
|||||

|
<a href="http://www.ammonit.ru/text/1327.htm#249459">http://www.ammonit.ru/text/1327.htm#249459</a> |
|
|||||
По моим наблюдениям как в "скученных" захоронениях (набитые раковинами конкреции или прослои), так и просто в слое (если раскапывать его фиксируя каждую, даже плохонькую, находку) количество микроконхов и соразмерной молоди макроконхов сопоставимо. Случаев исключительного или почти исключительного захоронения микроконхов, по крайней мере, со статистически достоверной выборкой, я что-то вот так сразу не припомню. При этом вспоминаются не самые редкие случаи захоронения (преимущественно конкреционного) исключительно или почти исключительно макроконхов. Но вот какой из факторов - этологический или тафономический - здесь играл роль сказать сложно. | ||||||
|
комментарий 1 уровня |
Комментарий создан 6 ноября 2015 года в 00:37:40 |
|||||

|
<a href="http://www.ammonit.ru/text/1327.htm#249472">http://www.ammonit.ru/text/1327.htm#249472</a> |
|
|||||
Могу пригласить тебя в Бронницы (Марково) - там в зоне A.alternoides амоебоцерасовых макроконхов крайне мало, в Михайлов на нижний оксфорд - там тоже кардиоцерасовые макроконхи очень редки по сравнению с микроконхами. В Москве и области на всех разрезах зоны fulgens многократный перевес в численности у микроконхов. Как я понимаю здесь - Долина р. Б. Зеленчук, р-н Скалистого хребта макроконхи тоже редки. Понятно, что это не везде так, например в Бронницах (Рыбаки) в зоне A.serratum макроконхов амоебоцерасов полно, ничуть не меньше, чем микроконхов. В Михаленино их и в альтерноидесе полно, не то что в Марково.
| ||||||
|
комментарий 2 уровня |
Комментарий создан 6 ноября 2015 года в 10:08:27 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#249477">http://www.ammonit.ru/text/1327.htm#249477</a> |
|
|||||
В Марково мы только по очень косвенным признакам можем утверждать, что найденные фрагмоконы принадлежат микроконхам... | ||||||
|
комментарий 3 уровня |
Комментарий создан 6 ноября 2015 года в 11:21:51 |
|||||

{kind=link}
|
<a href="http://www.ammonit.ru/text/1327.htm#249480">http://www.ammonit.ru/text/1327.htm#249480</a> |
|
|||||
А ты не обращал внимание, сколько там аммонитов с пиритизированными жилыми камерами? Конечно меньше, чем фрагмоконов, но все-таки много (они хорошо опознаются по одной продавленной стороне ЖК), плюс находки в глине, где жилую камеру видно, даже если взять нельзя.
| ||||||
|
комментарий 4 уровня |
Комментарий создан 6 ноября 2015 года в 12:23:20 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#249482">http://www.ammonit.ru/text/1327.htm#249482</a> |
|
|||||
Фрагмоконы целиком - да. Но у них нет разворота спирали, и мы не знаем, есть ли финальное сближение перегородок (надо кстати будет приполировать несколько характерных образцов...)
| ||||||
|
комментарий 5 уровня |
Комментарий создан 6 ноября 2015 года в 12:45:16 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#249483">http://www.ammonit.ru/text/1327.htm#249483</a> |
|
|||||
Как раз у микроконхов раскручивание выражено не всегда и слабее, чем у макроконхов. А вот сближение должно быть, тут действительно надо пришлифовывать.
| ||||||
|
комментарий 6 уровня |
Комментарий создан 6 ноября 2015 года в 12:53:55 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#249486">http://www.ammonit.ru/text/1327.htm#249486</a> |
|
|||||
Ну да, я и имел в виду, что и у взрослых микроконхов, и у молодых всяких-конхов раскручивания спирали нет.
| ||||||
|
комментарий 7 уровня |
Комментарий создан 6 ноября 2015 года в 13:01:01 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#249488">http://www.ammonit.ru/text/1327.htm#249488</a> |
|
|||||
В принципе может быть. Ведь с мелочью действительно непонятно, хотя надо бы и их насчет перегородок пришлифовать. | ||||||
|
комментарий 8 уровня |
Комментарий создан 6 ноября 2015 года в 13:05:07 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#249487">http://www.ammonit.ru/text/1327.htm#249487</a> |
|
|||||
Кстати по поводу раскручивания спирали у МАКРОконхов из Марково, там всё тоже как-то мутно. Иногда оно есть, иногда нет (хоть в обоих случаях есть сглаживание скульптуры). Но их слишком мало, чтобы пытаться делать какие-то выводы. | ||||||
|
комментарий 7 уровня |
Комментарий создан 6 ноября 2015 года в 13:03:44 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#249490">http://www.ammonit.ru/text/1327.htm#249490</a> |
|
|||||
И их мало, и более ранних кардиоцерасовых макроконхов мало, трудно сравнивать. Интереснее конечно понять куда делись макроконхи выше серратума, вот вопрос. | ||||||
|
комментарий 8 уровня |
Комментарий создан 6 ноября 2015 года в 13:11:42 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250112">http://www.ammonit.ru/text/1327.htm#250112</a> |
|
|||||
Насколько я понимаю, в Марково сбор аммонитов ведется главным образом путем отмывки, корректные данные можно получить только при детальном раскапывании и хорошем навыке различения макро- и микроконхов по молодым раковинам. Что касается зоны Fulgens, то я у этих ребят вообще сомневаюсь в возможности различения диморфов, по крайней мере, Миша так меня и не убедил. В в. байосе СЗ Кавказа я лично не копал, но СВ Кавказ (та же фация) показывает видимое отсутствие закономерности. А в н.байосе (зона Humphriesianum) у сферороцератид и стефаноцератид там явное равновесие. У сонниниид наблюдаются дисбалансы, но с учетом переотложенно-конкреционного захоронения вполне могут быть объяснимо снивелированы. | ||||||
|
комментарий 3 уровня |
Комментарий создан 10 ноября 2015 года в 12:18:31 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250113">http://www.ammonit.ru/text/1327.htm#250113</a> |
|
|||||
В Марково последние годы сбор аммонитов ведётся больше раскапыванием, с пиритом там довольно сконденсированный слой. Но если абстрагироваться от пирита, в вышележащих слоях с плоскими "плюшками" я как-то тоже не припоминаю обилия взрослых макроконхов кардиоцератид (при том, что ниже - в подзоне ilovaiskii - их довольно много). | ||||||
|
комментарий 4 уровня |
Комментарий создан 10 ноября 2015 года в 12:34:24 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250133">http://www.ammonit.ru/text/1327.htm#250133</a> |
|
|||||
А как Вы отличаете молодых макроконхов кардиоцератид от "взрослых макроконхов кардиоцератид" от по тому материалу? | ||||||
|
комментарий 5 уровня |
Комментарий создан 10 ноября 2015 года в 13:55:17 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250136">http://www.ammonit.ru/text/1327.htm#250136</a> |
|
|||||
Из предположения, что взрослые макроконхи крупные. Т.к. изредка там всё же попадаются крупные, с ослаблением скульптуры на ЖК, иногда с небольшим разворачиванием спирали. Конечно, это может быть просто другой вид, а макроконхи alternoides - мелкие, трудноотличимые от микроконхов. А как с этим обстоит дело в Михаленино? | ||||||
|
комментарий 6 уровня |
Комментарий создан 10 ноября 2015 года в 14:03:58 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250175">http://www.ammonit.ru/text/1327.htm#250175</a> |
|
|||||
Вот, Вы же всё понимаете. О какой теперь корретности данных идет речь. Да, и я о том! | ||||||
|
комментарий 7 уровня |
Комментарий создан 10 ноября 2015 года в 18:43:39 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250237">http://www.ammonit.ru/text/1327.htm#250237</a> |
|
|||||
Сохранившиеся предыдущие обороты рассыпавшихся макроконхов с гладкой ЖК вполне себе alternoides-овые: Фрагменты гладких макроконхов A.alternoides. | ||||||
|
комментарий 7 уровня |
Комментарий создан 11 ноября 2015 года в 00:19:45 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250240">http://www.ammonit.ru/text/1327.htm#250240</a> |
|
|||||
Вполне возможно, что гладкие амебоцерасы - это не "макроконхи", а всего лишь представители пострепродуктивного периода развития, которого достигали редкие индивидуумы. | ||||||
|
комментарий 1 уровня |
Комментарий создан 11 ноября 2015 года в 02:14:23 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250243">http://www.ammonit.ru/text/1327.htm#250243</a> |
|
|||||
А известны ли головоногие, активно растущие в пострепродуктивном периоде? | ||||||
|
комментарий 2 уровня |
Комментарий создан 11 ноября 2015 года в 07:46:43 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250250">http://www.ammonit.ru/text/1327.htm#250250</a> |
|
|||||
не стоит забывать, что аммоноидеи - специфическая группа головоногих и принцип актуализма в отношении их не всегда работает | ||||||
|
комментарий 3 уровня |
Комментарий создан 11 ноября 2015 года в 10:56:15 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250251">http://www.ammonit.ru/text/1327.htm#250251</a> |
|
|||||
Ну, так им можно что угодно приписать. На самом деле у современных головоногих нет никакого "старческого роста" и нет никаких оснований предполагать его у аммонитов.
| ||||||
|
комментарий 4 уровня |
Комментарий создан 11 ноября 2015 года в 11:16:02 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250254">http://www.ammonit.ru/text/1327.htm#250254</a> |
|
|||||
Я совсем не настаиваю. А какую роль Вы отводите стадии гладкой КЖК у амебоцерасов? Зачем она им была нужна?
| ||||||
|
комментарий 5 уровня |
Комментарий создан 11 ноября 2015 года в 11:46:20 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250263">http://www.ammonit.ru/text/1327.htm#250263</a> |
|
|||||
Однозначного ответа у меня нет. Но есть интересное исследование Chamberlain J.A., Westermann G.E.G. (1976) Hydrodynamic properties of cephalopod shell ornament. в котором показано, что коэффициент сопротивления при движении раковины в воде довольно неожиданным образом зависит от скульптуры - небольшие скульптурные элементы на небольших раковинах снижают сопротивление, но по мере роста раковины и увеличения размеров скульптуры позитивный эффект исчезает (авторы пишут "To be hydrodynamically optimal, small shells (diameter ≈ l0 cm) must have rough (sculptgred) surfaces, wheieas large shills (diameter ≈ 100 cm) require smooth surfaces."). Тут конечно не все так однозначно, имеет значение форма раковины и характер движения (рывок с места или длительное плавание). Но вполне возможно, что именно это частично объясняет сглаживание скульптуры. Правда, скульптура сглаживалась у макроконхов с разной формой раковины, в том числе у кадиконов вроде Rondiceras или Cadoceras, но тут может быть что один и тот же результат получался в силу разных причин, а может и для них гидродинамика все-таки была важна. Это же объясняет, почему мало менялась скульптура у дорсопланитид - с их колесовидной формой раковины такие гидродинамические тонкости явно не имели значения.
| ||||||
|
комментарий 6 уровня |
Комментарий создан 11 ноября 2015 года в 12:36:51 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250265">http://www.ammonit.ru/text/1327.htm#250265</a> |
|
|||||
вот, кстати, тоже вариант к вопросу "почему их так мало?".
| ||||||
|
комментарий 7 уровня |
Комментарий создан 11 ноября 2015 года в 12:47:09 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250268">http://www.ammonit.ru/text/1327.htm#250268</a> |
|
|||||
Да, так могло быть, но только в том случае, если реализация скульптуры зависела только от достигнутого размера (и фактически сам аммонит определял, когда пора ее менять). Если изначально было генетически заложено, что "девочка" будет большая, а значит с определенного момента развития (ее внутреннего гормонального развития) у нее генетически заложено изменение скульптуры - это уже другое дело.
| ||||||
|
комментарий 8 уровня |
Комментарий создан 11 ноября 2015 года в 13:32:04 |
|||||
{kind=link}
|
<a href="http://www.ammonit.ru/text/1327.htm#250282">http://www.ammonit.ru/text/1327.htm#250282</a> |
|
|||||
Возможно слишком консервативен, но по фото я бы сказал, что левый экземпляр тяготеет к A.glosense, а второй - к A.alternoides.
| ||||||
|
комментарий 9 уровня |
Комментарий создан 11 ноября 2015 года в 14:36:42 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250285">http://www.ammonit.ru/text/1327.htm#250285</a> |
|
|||||
Но ведь A.glosense это и есть макроконх альтерноидеса, просто этот вид описан по макроконховому фрагмокону. А по сути это тот же A.alternoides, не даром их так или иначе объединяют в один вид.
| ||||||
|
комментарий 10 уровня |
Комментарий создан 11 ноября 2015 года в 15:00:06 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250392">http://www.ammonit.ru/text/1327.htm#250392</a> |
|
|||||
Разве, что A.transitorium, судя по голотипу, не очень стремился к избавлению от ребер | ||||||
|
комментарий 11 уровня |
Комментарий создан 12 ноября 2015 года в 02:27:05 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250533">http://www.ammonit.ru/text/1327.htm#250533</a> |
|
|||||
К сожалению, книги Спэта у меня нет, но если речь о размере голотипа, у Мессежникова (1989) я нашел интересный момент:
| ||||||
|
комментарий 12 уровня |
Комментарий создан 13 ноября 2015 года в 13:47:06 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250652">http://www.ammonit.ru/text/1327.htm#250652</a> |
|
|||||
Книга Спэта (с юрассик.ру): http://ashipunov.info/jurassic/j/Spath1935.pdf
| ||||||
|
комментарий 13 уровня |
Комментарий создан 14 ноября 2015 года в 10:20:26 |
|||||
|
<a href="http://www.ammonit.ru/text/1327.htm#250653">http://www.ammonit.ru/text/1327.htm#250653</a> |
|
|||||
Спасибо за ссылку! Я ее там как-то проморгал...
| ||||||
|
комментарий 14 уровня |
Комментарий создан 14 ноября 2015 года в 11:00:16 |
|||||